 .Figura 1. Filamentos de
actina (Fonte: ALBERTS B.Fundamentos da Biologia Celular)
.Figura 1. Filamentos de
actina (Fonte: ALBERTS B.Fundamentos da Biologia Celular)"Trichomonas vaginalis e importância do seu citoesqueleto "
ANA BEATRIZ GUIMARÃES, CARLOS EDUARDO SAMPAIO GUEDES, FLÁVIO AUGUSTO RIBEIRO DE SOUZA, ILAI MORADILLO MELLO ALVES, LIA MIDORI MEYER NASCIMENTO & ZAFIRA EVELMA GURGELProfessor Orientador Solange Peixinho
INTRODUÇÃO
O citoesqueleto, mais especificamente os microfilamentos de actina são a arma secreta da Trichomonas vaginalis o agente etiológico causador da tricomoníase, uma D.S.T.(doença sexualmente transmissível) que provoca a infecção do aparelho genital do homem e da mulher. É através deles que a Trichomonas vaginalis tira seu disfarce de " boa samaritana" ao regredir seu flagelo e originar seus " temíveis " pseudópodos fixadores, mostrando sua " verdadeira face ", a de um parasito que é infeccioso e impiedoso.
Portanto o objetivo desse trabalho é abordar aspectos estruturais e moleculares dos microfilamentos de actina, associando-os a fisiologia da Trichomonas vaginalis, bem como a exposição de aspectos patológicos do parasito, as técnicas e os métodos utilizados neste tipo de abordagem e suas finalidades.
CITOESQUELETO E SUA FUNÇÃO
Nos eucariontes, a estrutura que lhes permite adotar uma variedade de formas e executar movimentos coordenados depende de uma complexa rede de filamentos protéicos denominado citoesqueleto. Esta rede é formada por três principais tipos de filamentos: os microtúbulos, os filamentos intermediários e os microfilamentos. Tais filamentos diferem entre si quanto à estrutura, dimensões e funções que desempenham na célula.
Os microtúbulos são constituídos de subunidades de tubulina agrupadas de tal forma a originar um tubo oco de 25 nm de diâmetro, desempenhando funções como transporte de organelas nas células, formação do fuso mitótico , fluxo de substâncias no interior da célula e formação dos cílios e flagelos – flagelos estes que promovem a locomoção da Trichomonas vaginalis quando esta não está parasitando.
Já os filamentos intermediários são formados por monômeros de muitas proteínas, possuem um diâmetro de 10 nm e apresentam diversas funções, dentre elas, a formação da lâmina nuclear que organiza e desorganiza a carioteca antes e depois da divisão, respectivamente.
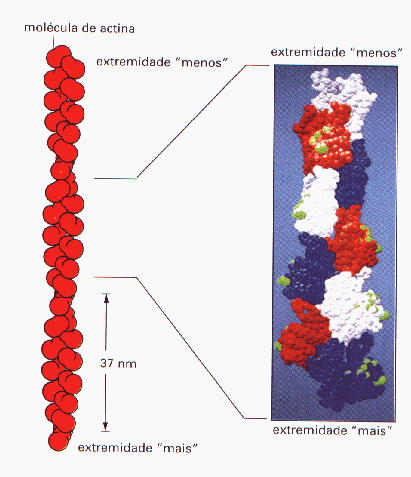
Além desses, existem ainda os microfilamentos de actina, que são alvos principais deste trabalho. Eles são constituídos de actina globular, ou g-actina, que fazem uma arrumação em hélice possuidora de uma extremidade "mais" (+) e uma extremidade "menos" (-). A actina tem, como papel, a sustentação de microvilosidades e outras projeções celulares, contração, ciclose e formação de pseudópodos que atuam na forma citotóxica da Trichomonas vaginalis, como será abordado mais adiante .
FILAMENTOS DE ACTINA
A actina é uma proteína bastante concentrada no citoesqueleto e presente em todas as células eucarióticas. Ela é importante para os movimentos celulares, podendo formar, nas células, estruturas lábeis ou estáveis.
Os filamentos de actina são filamentos que possuem duas extremidades diferentes - uma "mais" que possui um crescimento rápido e uma "menos", relativamente inerte (Fig.1). A polimerização in vitro da actina pura necessita de ATP e ions monovalentes ou divalentes de K ou Mg . Quando os ions são adicionados a monômeros de actina na presença de ATP, ocorre um aumento dos filamentos devido à polimerização rápida. A velocidade de polimerização é diferente em cada extremidade do filamento, sendo que, na extremidade "mais", ela é dez vezes mais rápida que na "menos".
A hidrólise de ATP é importante para a despolimerização dos filamentos, pois enfraquecerá as ligações no polímero. Este processo contribui para a troca rápida de subunidades existentes na célula. As moléculas de actina se polimerizam e despolimerizam no córtex das células, gerando microespículas e lamelipódios. Estas estruturas seriam geradas por uma polimerização local da actina nível da membrana plasmática, o que proporcionaria à membrana um impulso para frente sem se romper, gerando a formação de uma estrutura denominada pseudópodo que permite organismos, como a Trichomonas vaginalis, fixar-se ao hospedeiro
.Figura 1. Filamentos de
actina (Fonte: ALBERTS B.Fundamentos da Biologia Celular)
TRICHOMONAS VAGINALIS
Trichomonas vaginalis, pertencente à família Trichomonadidae, foi descrita em 1836, presente em uma paciente com vaginite e, tempos depois, encontrada na uretrite de um homem.
Protozoário parasita e flagelado, possui a capacidade de formar pseudópodos que servem para a captura de alimentos e para se fixar em partículas sólidas, porém estes pseudópodos não realizam movimentos amebóides. As Trichomonas vaginalis transitam da forma flagelada para amebóide/aderente a depender da disponibilidade de suprimentos alimentares, que são ingeridos por fagocitose através dos finos pseudópodos.
É no trato geniturinário do homem e da mulher que o parasito produz a infecção – ele não sobrevive fora do sistema urogenital. Sua reprodução ocorre por divisão binária longitudinal e não há formação de cistos. A fisiologia pode ser descrita como organismos anaeróbios facultativos; como fonte de energia, utiliza monossacarídeos, principalmente o glicogênio; possuem numerosas enzimas glicolíticas e são desprovidos de mitocôndrio e citocromo, portanto o ciclo de Krebs é incompleto.
A transmissão ocorre através da relação sexual – é uma doença venérea. Em raros casos, pode ser transmitida durante o parto ou com o uso de objetos de uso pessoal e íntimo.
Em geral, a infecção ocorre no trato urogenital e os sintomas e sinais se manifestam de maneira diferente nos homens e nas mulheres. Nas mulheres, provoca vaginite, com dores e dificuldades miccionais e nas relações sexuais; a vagina e a cérvix mostram pontos hemorrágicos. Nos homens, provoca uretrite. A infecção pode se dar na próstata, na bexiga, na vesícula seminal e nos testículos, causando algumas patogenicidades, como cistite, prostite, epididimite e outras. A infecção pode estar associada a uma diminuição do glicogênio e enfraquecimento da camada epitelial.
A Trichomonas vaginalis não é sensível a antibióticos, mas muitas cepas apresentam, atualmente, resistência ao metronidazol; a vacino-terapia é uma tentativa de tratamento da tricomoníase.
A TRICHOMONAS VAGINALIS E O CITOESQUELETO DE ACTINA
Muitos estudos sobre os Tricomonadídeos têm sido realizados nos últimos anos, a maioria deles devotados ao estudo do citoesqueleto e a sua relação com a citotoxidade de alguns membros do grupo. Nesses estudos, as espécies mais utilizadas são: a Tetratrichomonas gallinarum, Trichomitus batrachorum, Tritrichomonas foetus, Tritrichomonas tenax, Pentatrichomonas hominis, Trichomonas equi e a Trichomonas vaginalis, que é alvo da maioria dos estudos. Dela, os cientistas removem o citoesqueleto de actina estudam sua composição bioquímica e sua relação com a patogenicidade da forma amebóide , pois a sua forma flagelada não pode parasitar.
Na forma amebóide observou-se uma grande quantidade de actina-G e de filamentos de actina polimerizada (actina-F) demonstrando a sua importância na formação de pseudópodos, com os quais esta alimenta-se e fixa-se ao hospedeiro.
Esta transição da forma flagelada para amebóide não era muito bem explicada há alguns anos, este aliado ao fato de terem sido encontradas Trichomonsa vaginalis em mulheres que não apresentavam a tricomoniase levou muitos cientistas a acreditarem que esta fosse uma "boa samaritana", sendo acusada injustamente por este fato. Mas estudos mais aprofundados mostraram que a Trichomonas vaginalis realmente era a causadora da tricomoniase, e que ela "tira seu disfarce de boa samarita na" ao transitar da forma flagelada para amebóíde/aderente, consumindo o glicogênio da mucosa vaginal e enfraquecendo o tecido epitelial.
Nestas pesquisas foi demonstrado que a actina da Trichomonas vaginalis é muito parecida com a actina dos vertebrados, diferindo apenas em algumas isoformas comprovadas pela eletroforese bidimensional. Essa actina foi observada na forma amebóide com a utilização de immunoblotting, de anticorpos anti-actina e corantes fluorescentes em microscopia de imunofluorescência, eletroforese bidimensional e microscopia eletrônica por immunogold, além da ultracentrifugação .
Muitos pesquisadores atualmente baseados no estudo do citoesqueleto de actina, atribuem a citotoxidade da Trichomonas vaginalis a este citoesqueleto , que se apresenta em grande quantidade na sua forma amebóide. Ao formar os pseudópodos, a actina da Trchomonas vagi nalis polimeriza e despolimeriza, produzindo filamentos lábeis que dão forma ao citoplasma desta, demonstrando que, sem esse citoesqueleto ela não pode formar pseudópodos e conseqüentemente não pode parasitar e se alimentar, portanto "morre de fome".
Esses estudos são importantes para o desenvolvimento de medicamentos que atuem sobre o citoesqueleto de actina e tenham, conseqüentemente, mais eficiência que os medicamentos atualmente utilizados ( já que a Trichomonas vaginalis tem apresentado resistência ao metronidazol com já citado anteriormente), como a tricomicina, aminotrozol e nitrimidazina, abrindo campo inclusive para substâncias inibidoras de polimerização e estabilizadores de actina, como a citocalasina e a faloidina , respectivamente.
CONCLUSÃO
Foi observado no estudo que a proteína de actina é encontrada em todo o mundo eucariótico, já que está presente tanto nos tricomonadídeos quanto nas células humanas. Ela mostra-se altamente conservada, com exatamente 42 Kda em ambos os organismos, diferindo apenas em duas isoformas, ou seja, a actina é uma proteína fundamental para os organismos eucarióticos.
A função parasitária da Trichomonas vaginalis está intrinsecamente associada a aspectos funcionais e estruturais dos microfilamentos de actina, pois, com a regressão do flagelo e posterior polimerização da g-actina originando o filamento, proporciona a projeção da membrana. Essa projeção origina um pseudópodo fixador, o que permite à Trichomonas vaginalis parasitar o trato urogenital humano. Portanto, a finalidade do estudo do citoesqueleto consiste em elucidar uma provável técnica de combate à doença.
AGRADECIMENTOS
Nossos agradecimentos às professoras da UFBA., Ana Verena Madeira (Departamento de Zoologia, IBIO), Lúcia Perdiz (Departamento de Biologia, IBIO) e Luzimar Fernandez (Departamento de Bioquímica, ICS), pelo auxílio dado à nossa equipe.
FONTES BIBLIOGRAFICAS USADAS:
BRUCE, Alberts el al. Molecular biology of the cell. New York & London. Garland, 1983. 1146 p.
LENINGER, Albert L. Princípios de Bioquímica, São Paulo, SAVIER, 1984. 725 p.
NEVES, David Pereira. Parasitologia humana. 6 ed. Rio Janeiro, Atheneu, 1986. 665 p.
PESSOA , Samuel B. MARTINS, Amilcar V. Parasitologia médica. 11 ed. Rio de Janeiro: Guanabara, 1988. 872 p.
STRYER, Lubert. Bioquímica. 4 ed. Pio de Janeiro: Guanabara, 1996. 100 p.
ADDIS, M.F. et al. Cloning and molecular characterization of a cDNA clone cod for Trichomona vaginalis alpha- actinin and intracellular localization of the protein. National Library of Medicine. Infect Immun 1998 Oct
BRICHEUX, G. et al. Characterization, clonin and immunolocalization of a coron homologue in Trichomons vaginalis. National Library of Medicine. Eur J Cell 2000 Jun
BRICHEUX, G. et al. Evidence for a uncommon alpha-actin protein in Trichomonas vaginalis. National Library of Medicine. Mol Biochem Parisitol 1998 Sep
BRICHEUX, G. Brugerolle , G.Molecular clonin of actin genes in Trichomonas vaginalis na phylogeny inferred from actin sequences. National Library of Medicine. FEMS Microbiol Lett 1997 Aug.
BRUGEROLLE, G.; BRICHEUX,G.; COFFE,G. Actin cytoskeleton demonstration in Trichomonas vaginalis and in other trichomonads. Biol. Cell.,1996, 88, 29-36.
FIORI, PL. et al. The flagellate parasite Trichomonas vaginalis: new insights cytopathogenicity. National Library of Medicine. Microbes Infect 1999 Feb
GUILLEN N. et al. Cytoskeletal actin and its associated proteins. Some example Protista. National Library of Medicine. Parasite 1998 Jun.
SHAIO MF, et al. A novel neutrophil-activating factor realeased by Trichomonas vaginalis. National Library of Medicine. Infect Immum 1992 Nov.
Simões, MLPC.; Tavares-Souza, MB. Manual-Guia sobre eletroforese de proteínas e suas aplicações. ENCONTRO DE GENÉTICA DO NORDESTE. 13, s.d
GLOSSÁRIO
Citocromo: proteína colorida, contendo um grupo heme, que transfere elétrons durante a respiração celular e fotossíntese.
Eletroforese Bidimensional: técnica utilizada para separar e identificar proteínas presentes em uma mistura quando submetidas a um campo elétrico. É composta de dois procedimentos, o primeiro baseado n o pH característico das proteínas, e o pI (ponto isoelétrico), e o segundo baseado no peso molecular e na carga elétrica específica das mesmas. específica das mesmas.
Glicogênio: polissacarideo composto exclusivamente por unidades de glicose, utilizada para armazenar energia em células animais
Immunoblotting: técnica que permite identificar uma molécula especifica entre diversas, com a utilização de um anticorpo marcado com um isótopo radioativo, que ao se ligar a essa molécula específica, permite que a presença da mesma seja detectada.
Imunofluorescência: técnica usada para identificar uma molécula especifica, através da ação de um anticorpo, marcado com um corante fluorescente, que se ligará a essa molécula e permitirá a detecção da mesma.
Imunogold: técnica empregada para identificar ma molécula específica, entre diversas, através da utilização de um anticorpo com esferas do ouro coloidal. Ele se ligará à molécula específica fazendo com que a mesma possa ser identificada.
Isoforma: uma das várias formas da mesma proteína que possui alguma diferença na seqüência de aminoácidos
ligar a essa molécula específica; permite que a presença da mesma seja detectada.
Pseudópodo: grande protrusão da superfície celular formada pelas células amebóides. À medida que estas vão se movimentando. Mais genericamente, qualquer extensão dinâmica, rica em actina, na superfície da célula animal.
Ultracentrifugação: procedimento que permite o isolamento de proteínas e organelas, baseada na diferença de tamanho e densidade entre as mesmas que ao serem giradas em torno de m eixo a velocidades altas separam-se em um gradiente de densidade.